您當前的位置:檢測資訊 > 實驗管理
嘉峪檢測網 2025-03-19 08:24
1. 背景介紹
蛋白質是生命活動的核心分子,其功能由其三維結構決定。蛋白質的結構可分為四個層次:一級結構(氨基酸序列)、二級結構(α螺旋、β折疊等局部構象)、三級結構(單個多肽鏈的全局折疊)和四級結構(多亞基的組裝)。
例如,酶的催化活性依賴于活性中心的精確三維排布,膜蛋白的信號轉導功能與其跨膜結構域的構象變化密切相關。
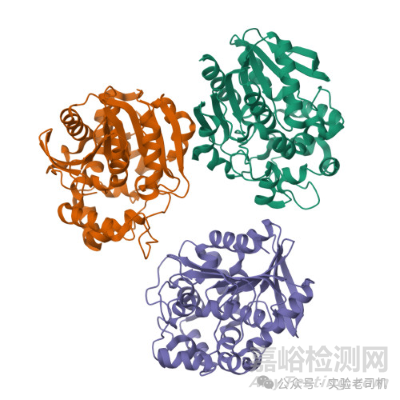
(圖片來源:https://www.rcsb.org/#Category-analyze)
研究蛋白質結構不僅能揭示其作用機制,還為藥物設計、疾病治療和合成生物學提供基礎。然而,蛋白質的尺寸通常在納米級別,傳統光學顯微鏡無法觀測,因此需要借助高分辨率技術解析其原子級結構。
X射線晶體學(XRD)和冷凍電子顯微鏡(Cryo-EM)是當前兩種最主流的蛋白質結構解析技術。
2. 方法學比較
● X射線晶體學(XRD)
X射線晶體學的發展始于20世紀初,英國物理學家William Henry Bragg與其子William Lawrence Bragg開創了X射線晶體學領域。他們提出的布拉格定律揭示了X射線與晶體原子排列的定量關系,為物質結構分析奠定了數學基礎。
這項技術的影響之深遠,正如奧地利化學家馬克斯·佩魯茨所言:“27項諾貝爾獎不可或缺的配方”——截至20世紀末,X射線晶體學直接推動了27項諾貝爾獎的誕生。1953年,英國科學家羅莎琳德·富蘭克林(Rosalind Franklin)利用X射線晶體學拍攝了DNA的B型衍射圖(“照片51號”)。
圖中的“X”形衍射斑直接提示了螺旋結構的對稱性——每圈10個堿基對,螺距34 Å。盡管富蘭克林因早逝未能獲得諾貝爾獎,但她的這份圖像幫助詹姆斯·沃森和弗朗西斯·克里克創建了DNA 分子的雙螺旋結構模型,成為了生物學發展的一座里程碑。
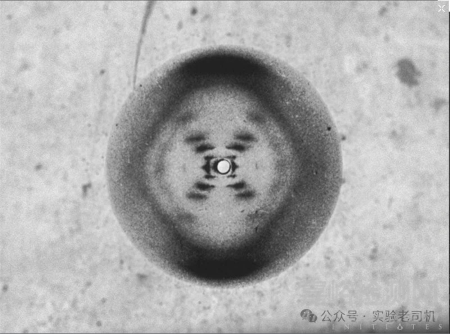
(“照片51號”;圖片來源:https://assassinscreed.fandom.com/zh/wiki)
1958年,科學家在首個成像蛋白質—肌紅蛋白內發現了不規則的褶皺,首次利用X射線晶體學解析了其結構,標志著X射線晶體學在蛋白質結構分析領域的重大突破。
該技術的核心原理是利用X射線與蛋白質晶體中原子的相互作用:當X射線穿過高度有序的晶體時,晶格中的原子會使X射線發生衍射,形成特定的衍射圖案。
通過分析這些衍射點的位置和強度,結合數學方法(如傅里葉變換),可計算出蛋白質的三維電子密度圖,進而構建原子模型。
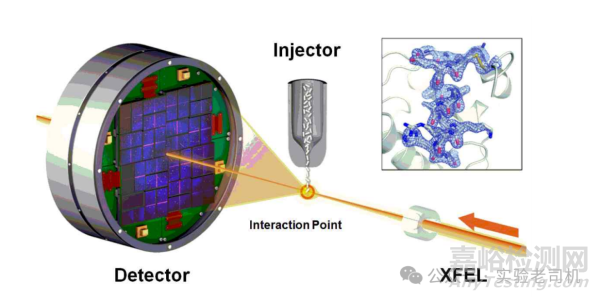
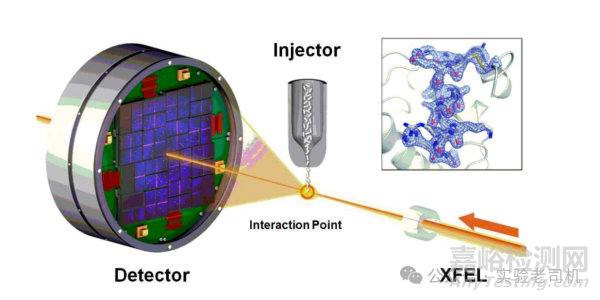
(圖片來源:https://images.app.goo.gl/yfVo1LwXBFu5Dsxq5)
然而,該技術面臨兩大挑戰:蛋白質結晶和相位問題。許多蛋白質(尤其是膜蛋白或柔性蛋白)難以形成高質量晶體。例如,G蛋白偶聯受體(GPCR)因跨膜區的不穩定性,直到2004年通過引入抗體片段穩定構象才被成功解析。
相位問題則源于X射線衍射僅能記錄振幅信息而丟失相位(波的相對位置),需借助分子置換法或引入重金屬原子(如硒代甲硫氨酸)來間接解決。
技術的進步顯著提升了X射線晶體學的應用范圍。同步輻射光源提供了高強度X射線,使數據采集速度更快、分辨率更高。X射線自由電子激光(XFEL)甚至能捕捉飛秒級動態過程,例如光系統II分解水分子時的瞬時中間態。
● 冷凍電子顯微鏡(Cryo-EM)
冷凍電子顯微鏡的突破性進展始于2010年代,被稱為“分辨率革命”。與X射線晶體學不同,Cryo-EM無需結晶,可直接分析溶液中的蛋白質樣品。
其關鍵步驟包括:將蛋白質樣品快速浸入-196℃的液乙烷中,使其在毫秒內凍結成無定形的玻璃態冰,避免冰晶形成(冰晶會擠壓樣品導致結構畸變)。玻璃態冰中的水分子呈無序狀態,包裹生物分子并維持其天然構象。
隨后用高能電子束穿透樣品并記錄散射電子的信號。通過采集數萬至數百萬張二維投影圖像,利用計算機算法(如單顆粒分析)對隨機取向的顆粒進行三維重構。
(圖片來源:NobelPrize.org)
Cryo-EM技術的核心優勢在于解析柔性或超大復合體的能力。例如,核糖體和病毒衣殼(如乙肝病毒)的精細結構均通過Cryo-EM得以揭示。
此外,該技術能捕捉同一蛋白的多種構象狀態。在新冠病毒研究中,Cryo-EM解析了刺突蛋白的“開放”(結合宿主受體ACE2)和“閉合”(免疫逃逸)構象,為疫苗設計提供了關鍵依據。
Cryo-EM技術瓶頸的突破主要依賴于硬件和算法的革新。直接電子探測器(DED)顯著提升了圖像信噪比,而深度學習算法(如cryoSPARC)實現了高效的數據處理和分類。
目前,Cryo-EM的最佳分辨率已接近2 Å,與X射線晶體學相當,但對樣品的純度、濃度和均一性仍有較高要求。
3. 小結
X射線晶體學和冷凍電子顯微鏡是研究結構生物學的兩大支柱,兩種技術并非相互替代,而是形成互補。
X射線晶體學在原子級分辨率上具有優勢,以原子精度揭示靜態細節,尤其適合小分子量、剛性且可結晶的蛋白質。而Cryo-EM更擅長處理超大復合體(如核孔復合物)或動態過程(如分子伴侶輔助的蛋白質折疊),以動態視角捕捉構象變化。
二者共同推動了蛋白質功能機制的解析。隨著技術進步,XRD和Cryo-EM技術正與其他方法深度融合,人工智能(如AlphaFold)的介入進一步加速了結構預測與實驗驗證的迭代。
這些發展將推動結構生物學從單一分子解析邁向復雜系統的動態研究,最終實現“從原子到細胞”的全尺度生命理解。
4. 參考文獻
馬禮敦. X射線晶體學的百年輝煌 [J]. 物理學進展, 2014, 34(2): 47–117. doi: 10.13725/j.cnki.pip.2014.02.001
https://digitalpaper.stdaily.com/http_www.kjrb.com/kjrb/images/2014-03/30/02/DefPub2014033002.pdf
Palczewski, K., Kumasaka, T., Hori, T., Behnke, C. A., Motoshima, H., Fox, B. A., ... & Miyano, M. (2000). Crystal structure of rhodopsin: A G protein-coupled receptor. Science, 289(5480), 739-745.
Rasmussen, S. G., Choi, H. J., Rosenbaum, D. M., Kobilka, T. S., Thian, F. S., Edwards, P. C., ... & Kobilka, B. K. (2007). Crystal structure of the human β2 adrenergic G protein-coupled receptor. Nature, 450(7168), 383-387.
Rasmussen, S. G., DeVree, B. T., Zou, Y., Kruse, A. C., Chung, K. Y., Kobilka, T. S., ... & Kobilka, B. K. (2011). Structure of a nanobody-stabilized active state of the β2 adrenoceptor. Nature, 477(7366), 549-555.
McCoy, A. J. (2007). Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallographica Section D: Biological Crystallography, 63(1), 32-41.
LI Zhifei, GAO Ning. Cryo-EM Method for High-Resolution Structure Determination: A Brief Introduction to 2017 Nobel Prize in Chemistry. University Chemistry[J], 2018, 33(1): 1-6 doi:10.3866/PKU.DXHX201711035

來源:實驗老司機


